The Harmful Immunologic Consequences of
Vaccinating Children Against COVID
A Brief Review
Open Letter to Parents and Pediatricians—Part IV
By Robert Rennebohm, MD
September 10, 2022

Introduction:
Since March 2021 Dr. Geert Vanden Bossche has been sharing his scientific concerns about the COVID mass vaccination campaign. [1-9] Initially, he warned that the COVID mass vaccination campaign would result in a prolonged series of successive new dominant variants, each being more infectious than its predecessor. [1, 2] That is exactly what has happened. He also warned that eventually a more virulent variant would appear. That, too, is now becoming apparent. [10, 11] He has warned that continuation of the mass vaccination campaign will only prolong the pandemic and make it more dangerous. That is proving to be the case.
His concerns have been based on his deep and broad study of immunology, virology, vaccinology, molecular biology, and evolutionary biology. His understandings of how COVID vaccines cause problems at both the individual and population levels are based on a combination of: extensive review of sound peer-reviewed scientific publications; his own decades of experience and study of vaccinology; and his synthesis and scientific judgment regarding what is known and what is probable. As stated above, his predictions and scientific judgments have repeatedly squared with subsequent realities.
He has generously and altruistically shared his concerns and understandings [9] and has repeatedly asked the promoters of the mass vaccination campaign to engage in respectful, constructive, altruistic scientific dialogue.
Dr. Vanden Bossche’s most recently expressed concern is that COVID vaccination of young children will interfere with the foundational education of their innate immune system—particularly the education of Natural Killer (NK) cells— and, thereby, render such children less able to successfully manage SARS-CoV-2, less able to handle other infections, and more prone to autoimmune disease. [3-6]
Dr. Vanden Bossche’s critically important scientific concerns must be thoroughly and honestly addressed before continuing the COVID mass vaccination campaign. Unfortunately, the promoters of the COVID mass vaccination campaign have not honored Dr. Vanden Bossche’s concerns. Instead, they have stifled scientific dialogue and increased their efforts to vaccinate as many people as possible—including, now, infants and toddlers—despite inadequate study of the safety and efficacy of the vaccines.
The American Academy of Pediatrics (AAP) and most pediatricians are either unaware of Dr. Vanden Bossche’s concerns, doubt that Dr. Vanden Bossche’s understandings are accurate, or have otherwise chosen not to speak up about his concerns. Pediatricians tend to be kind, humble, respectful, cooperative individuals. In their humility, most pediatricians probably think their knowledge of immunology and vaccinology is not sufficiently deep to justify their challenging the prevailing COVID narrative and its mass vaccination campaign. Most pediatricians have preferred to trust and defer to the judgment, knowledge, and recommendations of the AAP and the CDC. Even if they have some reservations about the safety of the COVID vaccines, most pediatricians have been too busy to do their own critical examination of the science involved.
As a retired pediatrician who has spent decades studying and treating children with autoimmune disorders, I have had the background and the time to perform the homework that has been difficult for practicing pediatricians to do. The results of that homework are shared in a series of three Open Letters to Parents and Pediatricians. [12-15] As a pediatrician and pediatric rheumatologist, I feel an obligation to help bring Dr. Vanden Bossche’s critically important concerns and understandings to the attention of parents and pediatricians. Children and their parents deserve to have pediatricians speak up regarding these issues.
In this article I do my best to explain to parents and pediatricians why Dr. Vanden Bossche strongly feels that, on an immunologic basis alone, children should not be given any of the currently available COVID vaccines. In the tradition of healthy scientific dialogue, I welcome scientifically-sound counter-arguments to these concerns and understandings (as does Dr. Vanden Bossche).
This article adds to information already provided in the three previous Open Letters to Parents and Pediatricians [12-15].

Antibodies (Abs) induced by COVID vaccines outcompete and sideline the young child’s innate (natural) antibodies and, thereby, interfere with viral clearance and the foundational education of a young child’s NK (Natural Killer) cells:
The immune system is comprised of two major branches—the innate immune system and the adaptive immune system. (See Table below.)

The innate immune system is the “first responder” and provides a rapid, powerful, relatively non-specific response. The adaptive immune system reacts more slowly, provides a more specific and precise response, and has potential for excellent memory. The innate and adaptive branches work collaboratively and synergistically to protect us from infection. Often, the innate immune system is able to eradicate infection without needing to recruit the adaptive immune system.
During early childhood (particularly during infancy and toddlerhood), the innate immune system provides an abundance of invaluable innate antibodies (natural antibodies). [16-25] During the rest of childhood and throughout adulthood, innate antibodies are still available and still play an important role but are not as abundantly involved.
Innate antibodies are purposefully non-specific, or polyspecific, as opposed to narrowly antigen-specific (specific for a single virus or other pathogen) and can quickly and effectively neutralize a broad range of viruses (as well as other infectious agents). This neutralization (of a virus, for example) prevents the virus from entering (infecting) cells and, thereby, minimizes or even fully prevents illness. [3-6, 12-15]
If/when innate Abs are overwhelmed (e.g., by a particularly large viral load and/or a virus that is particularly infectious and easily enters cells) they may not be able to prevent such a virus from entering and infecting cells. In anticipation of that problem, innate Abs educate NK cells (Natural Killer cells) so that those NK cells can recognize and kill infected cells (thereby killing the virus within those infected cells and preventing progression of the infection). [3-6, 12-15, 26-28 ] NK cells do this by recognizing foreign “patterns” that appear on the surface of infected cells. When they see these foreign “patterns,” they kill the infected cell. [3, 4, 6, 13]
[NK cells are a type of cytotoxic (or cytolytic) lymphocyte. They are part of the innate immune system. They are different from cytotoxic (cytolytic) T cells, which are part of the adaptive immune system.]
In order for Innate Abs to educate the NK cells, the innate Abs must, first, interact with the virus—i.e. bind to the virus. [3, 4, 6, 13, 14] If innate Abs are unable to bind to viruses, the innate Abs cannot educate the NK cells.
The innate Abs not only educate the NK cells regarding how to recognize and kill infected cells, but they also teach the NK cells how to avoid inappropriately attacking and killing healthy cells. In other words, the innate Abs teach the NK cells how to distinguish “self” from “non-self.” We want our NK cells to attack non-self (infected cells that have foreign patterns on their surface, or cells that have become cancerous) but to know how to leave healthy cells (“self”) alone. [26-32]
Viruses are tricky, because they can decorate themselves with glycosylated patterns (sugar-coated peptide patterns) that mimic the normal glycosylation patterns that are present on the surface of healthy cells. In this way, viruses, with their “self-mimicking” glycosylation patterns (patterns that mimic normal glycosylating patterns), can make it difficult for the NK cells to accurately distinguish between “self” patterns and “self-mimicking foreign” patterns. Fortunately, innate Abs are very good at educating NK cells to accurately make this distinction.
Key Point: The above critically important education of NK cells (by innate Abs)—this foundational education of NK cells— occurs primarily during early childhood, which is why young children are purposefully blessed with a unique abundance of innate Abs. This best opportunity for optimal foundational education of NK cells occurs only once in a child’s lifetime—namely during early childhood. It is critically important to avoid interfering with this precious opportunity. Subsequently, innate Abs and NK cells will still be available (to older children and to adults), but if the NK cells missed out on optimal foundational education during early childhood, those NK cells will be less competent during the rest of life—less able to quickly and accurately recognize infected cells, less able to avoid attacking healthy cells, and less able to undergo further training/fine-tuning. [3, 4, 6, 13]
So, we should certainly avoid interfering with the foundational education of NK cells—which primarily occurs during early childhood.
But a central concern of Dr. Vanden Bossche is that such interference is exactly what happens when a young child is vaccinated against COVID with any of the available COVID vaccines. [3, 4, 6, 13] My understanding of this concern is shared below:
The COVID vaccines teach the adaptive arm of the immune system to make two types of antibodies to the spike protein of SARS-CoV-2—neutralizing Abs and non-neutralizing Abs. (See APPENDIX OF MEDICAL ILLUSTRATIONS at the end of this article, after the References.) Initially, the neutralizing Abs worked modestly well to neutralize the spike protein of the original virus (Wuhan strain) and, thereby, keep the virus from easily entering cells. However, the virus has since learned how to escape that neutralizing effect, and this has rendered these neutralizing Abs inadequately effective, outdated. [3, 4, 6, 13, ] We will get back to this problem in a minute, but first let’s explain how these vaccinal Abs interfere with the foundational education of a young child’s NK cells.
Both the neutralizing vaccinal Abs and (to a lesser extent) the non-neutralizing vaccinal Abs are capable of binding tightly to the virus (the SARS-CoV-2 virus in the case of the current pandemic). Innate Abs (natural Abs) bind only loosely to viruses. In vaccinated individuals the vaccinal Abs, therefore, outcompete the innate Abs for binding sites on the virus. This results in the innate Abs not being able to bind to the virus. The innate Abs are sidelined, unable to interact with the virus, unable to participate in the immune response.
As mentioned earlier, innate Abs need to bind to viruses in order to provide the foundational education of the young child’s NK cells. If the vaccinal Abs are preventing this binding, the innate Abs are unable to interact with the virus and unable to educate the NK cells. The foundational education of NK cells is, thereby, suspended, and during the current pandemic this suspension has been prolonged, such that young children (if vaccinated) are at high risk of missing most or all of their precious once-in-a-lifetime optimal opportunity to foundationally educate their NK cells.
Dr. Vanden Bossche’s concern is that this vaccinal interference with the foundational education of a young child’s NK cells could have profound consequences. [3, 4, 6, 13] COVID vaccination of a young child may render those children less able to clear SARS-CoV-2 infection. Because they have been outcompeted and sidelined, the innate Abs are unable to neutralize the virus, which is what they would normally do. Also, because they have been sidelined, the innate Abs are unable to provide the much-needed foundational education of NK cells. Because both the innate Abs and the NK cells are sidelined the child must now rely primarily on the vaccinal antibodies to fight the infection.
This leads to discussion of a further problem. The vaccinal neutralizing Abs, which initially worked modestly well to neutralize the virus (Wuhan strain), are no longer working well (because the virus has learned how to escape these neutralizing Abs). [33-41] As those neutralizing Abs have become far less neutralizing, the vaccinal non-neutralizing Abs have become more dominant on the scene. These non-neutralizing antibodies have two major effects. One is an infection-enhancing effect. [42-43] These non-neutralizing Abs bind to the N-terminal domain (NTD) of the spike protein. This binding causes a conformational change in the spike protein that facilitates the ability of the spike protein to attach to the ACE-2 receptor in host cells, thus facilitating entry of the virus into human cells. This is a form of antibody-dependent enhancement of infection (ADEI). (See APPENDIX OF MEDICAL ILLUSTRATIONS, particularly Figures 16, 21, and 22.) So, the vaccinal Abs not only sideline the innate Abs (and, thereby, the NK cells that depend on the innate Abs), they increase the infectiousness of the virus and, thereby, make the vaccinated person more susceptible to infection.
The second effect of the non-neutralizing vaccinal Abs is that they are currently (but not for long) providing some protection from severe disease. They are doing this by binding to virus that has been absorbed onto dendritic cells. [2-6, 9, 44, 45] Normally, those dendritic cells carry the virus from the upper respiratory tract (URT) down to the lower respiratory tract (LRT) where the virus is then released and can infect cells in the LRT, causing severe disease in the lungs. By binding to the virus absorbed onto dendritic cells, the non-neutralizing Abs inhibit this release of virus in the LRT. This represents a beneficial “virulence-inhibiting” effect of the non-neutralizing Abs. This virulence-inhibiting effect is what is thought to be largely responsible for “prevention of severe disease” in vaccinated people. However, it is only a matter of time before the virus, inevitably, learns how to escape this virulence-inhibiting effect of the non-neutralizing Abs—just as the virus long ago figured out how to escape the neutralizing effect of the vaccinal Abs.
Once the virus learns how to escape the virulence-inhibiting effect of the non-neutralizing Abs, the vaccinal Abs will provide neither a beneficial neutralizing effect, nor a beneficial virulence-inhibiting effect. Worse, the non-neutralizing Abs will continue their infection-enhancing effect (i.e., facilitating entry of virus into cells, thereby enhancing infection), rendering the vaccinated person more susceptible to infection. Furthermore, the vaccinated person will not have innate Abs and NK cells to help them, because the innate Abs have been outcompeted and sidelined by the non-neutralizing Abs. All of these unfortunate consequences of COVID vaccination will more profoundly affect the COVID vaccinated young child but will also apply to all COVID vaccinated people.
Let’s return to discussion of what happens when the foundational education of a young child’s NK cells is disrupted for a prolonged period of time (because of the sidelining of innate Abs by COVID vaccinal Abs). Dr. Vanden Bossche’s concern is that this disruption does not just interfere with the education of NK cells regarding how to recognize and kill cells that have been infected by SARS-CoV-2 (which is a glycosylated virus). This disruption also interferes with the education of NK cells regarding how to recognize and kill other glycosylated viruses (e.g., influenza virus, RSV, common ordinary coronaviruses, rotavirus, measles, mumps, rubella, varicella, dengue, Ebola, avian flu, smallpox, monkeypox, e.g.). [4, 6, 31] This is because those other glycosylated viruses are decorated with patterns that have segments that are identical to segments within the SARS-CoV-2 virus. When the NK cell is not educated to recognize and attack SARS-CoV-2 (and, instead, learns to ignore and tolerate the glycosylation pattern on SARS-CoV-2) those NK cells also learn to ignore and tolerate the similar glycosylation patterns on many other glycosylated viruses. In this way, the COVID vaccines not only render the vaccinated person’s NK cells less able to recognize and kill SARS-CoV-2, but also less able to recognize and kill other glycosylated viruses.
Also, when the foundational education of a young child’s NK cells is disrupted, those NK cells do not become properly educated to distinguish between “self” and “non-self.” This renders the vaccinated child more susceptible to accidentally killing normal healthy cells—i.e. it puts them at risk of developing autoimmune diseases. [4, 6, 29]
Likewise, when the foundational education of a young child’s NK cells is disrupted, those NK cells do not become properly educated to recognize and kill early cancer cells. This predisposes the COVID vaccinated child to malignancy. [4, 6]
Another potential consequence of the COVID vaccines is “immune exhaustion.” [7, 9] Vaccinated people (of all ages) are now repeatedly becoming re-infected with SARS-CoV-2. This is because the COVID vaccines are not neutralizing the virus, the non-neutralizing Abs are actually facilitating infection of cells, and large amounts of the very infectious virus are now circulating throughout highly vaccinated communities, making it highly likely that vaccinated people will become re-infected, over and over. And each re-infection (as well as each booster shot) results in an increase in the quantity of non-neutralizing infection-enhancing vaccinal Abs.
When vaccinated people become reinfected (which is now happening frequently, for the above reasons): they are unable to rely on innate Abs (which have become sidelined) to directly neutralize and help clear the virus; they are unable to rely on NK cells (which have also become sidelined) to kill virus-infected cells; their vaccinal neutralizing Abs are no longer neutralizing (and are, therefore, ineffective); their vaccinal non-neutralizing Abs are facilitating entry of virus into cells (i.e. enhancing infection); and the once-beneficial virulence-inhibiting effect of the non-neutralizing vaccinal antibodies is about to disappear. When the immune system is having difficulty handling infection, it activates NK-CTL (NK-like Cytotoxic T Lymphocytes, which are a subset of CD8+ cytolytic T cells—i.e. part of the adaptive immune system) to help kill infected cells. [46] When a vaccinated person becomes repeatedly re-infected, thereby necessitating repeated activation of large amounts of NK-CTL, key components of the immune system become “exhausted,” or depleted. This is what is meant by “immune exhaustion.”
Immune exhaustion then leads to the vaccinated person being less able to handle not only SARS-CoV-2 and other glycosylated viruses that cause acute infection, but also other latent infections, including EBV, CMV, herpes virus, even TB. This immune exhaustion also renders the immune system more prone to autoimmune mistakes and adversely affects the immune system’s cancer surveillance capabilities—i.e., its ability to recognize and kill early malignancies.
Another problem with COVID vaccination is loss of flexibility, due to “original antigenic sin.” [47, 48] The COVID vaccines train (“prime,” “irreversibly program”) the adaptive immune system to produce precise antigen-specific antibodies (e.g., antibodies to the Wuhan spike protein) and then recall/resume production of those same antibodies whenever the vaccinated person is exposed to the virus in the future. Both the original neutralizing Abs and the original non-neutralizing Abs are recalled whenever the vaccinated person receives a booster shot or is exposed to current or future SARS-CoV-2 variants or receives the new bivalent COVID vaccine. Unfortunately, these recalled original vaccinal Abs are no longer able to neutralize the virus (because the virus has become resistant to these original neutralizing Abs). Worse, the recalled original vaccinal Abs (against the Wuhan spike protein) are likely to interfere with the production and function of new neutralizing Abs to new variants. This is what is meant by “original antigenic sin.” In contrast, the normal immune system is very flexible—it does not become programmed to forever respond only in its original way.
Whereas COVID vaccination of a young child exposes them to the above risks, a young child who is left unvaccinated against COVID will benefit in the following ways:
- The child’s abundant innate Abs will be free to bind to and neutralize the virus, including new variants—thus, contributing to clearance of the virus. (The child’s innate Abs will not be outcompeted and sidelined by vaccinal Abs.)
- The innate Abs will be free to provide a proper foundational education of NK cells—teaching the NK cells how to recognize and kill infected cells and cancerous cells, while also learning how to avoid killing normal healthy cells.
- The above healthy unimpaired innate immune system (innate Abs and NK cells, etc.) will be of tremendous help in containing and clearing the infection—so much so that the adaptive immune system might not need to be recruited.
- If needed, the child’s adaptive immune system will be able to produce new variant-specific (e.g. Omicron 2.75) Abs—unlike the vaccinated child whose immune system has been “primed” (“irreversibly programmed”) to respond to new variants with the same old outdated anti-Wuhan strain Abs (“original antigenic sin”), which interfere with successful development and function of new, updated antibodies. It is important to realize that once a person has become vaccinated, they cannot become de-vaccinated. Vaccination is irreversible.
- The unvaccinated child will not be burdened with “infection-enhancing” non-neutralizing vaccinal antibodies that render vaccinees more susceptible to infection.
- Significant natural infection with SARS-CoV-2 will result in long lasting sterilizing immunity, and such a child will be contributing to herd immunity. In contrast, vaccination does not result in sterilizing immunity and will not contribute to herd immunity. [14]
- The unvaccinated child will be able to respond normally to other glycosylated viruses.
- The unvaccinated child’s immune system will not become “exhausted.”
- The young child who is left unvaccinated will be in the best position of anyone to adequately handle any SARS-CoV-2 variant that comes along—even soon-to-arrive variants that are extremely infectious and severely virulent. In contrast, young children (especially 6-month-old children) who have received the COVID vaccine will be in the worst position of all people.
- However, when a more virulent variant appears on the scene (which Dr. Vanden Bossche thinks is inevitable and will occur soon), even unvaccinated children, despite being better off than vaccinated children, will be at greater risk of severe illness and hospitalization than has been the case to date—especially if the mass vaccination campaign is continued. This increased frequency of severe illness in unvaccinated children will not be because they are unvaccinated—it will be because of the increased infectiousness and increased virulence of the new variant(s). It is essential to understand that, at the population level, both the increased infectiousness and the increased virulence are direct results of the mis-guided COVID mass vaccination campaign. Neither would have developed in the absence of the mass vaccination campaign.
Because of the above-explained immunological concerns, young children should not be given the COVID vaccine. This applies to all COVID vaccines. It will also apply to the new bivalent “omicron-updated” mRNA vaccine that, unfortunately, is about to be rolled out without adequate testing or thoughtfulness. [8]
For similar reasons, older children and adolescents (and adults for that matter) should not be given the COVID vaccines: In older children, adolescents, and adults the vaccinal interference with the foundational education of NK cells is not an issue (because that education was likely accomplished during their early childhood), but they need to be able to use their innate Abs to help neutralize the virus and to provide further fine-tuning and training (ongoing, continuing education) of their NK cells. Except for the foundational education issue, the rest of the concerns mentioned regarding young children also apply to older children, adolescents, and adults.
For the above immunologic reasons alone, the COVID mass vaccination campaign should be halted— for all of Humanity, especially for young children—at least until these concerns have been thoroughly addressed.
Please note that I have not even mentioned the multiple other reasons that would (each, alone) justify an immediate halt to the mass vaccination campaign—e.g. the extremely inadequate study of both the short and long term safety of the COVID vaccines, both prior to and subsequent to approval of the vaccines; the extremely worrisome adverse effects of the COVID vaccines that have been reported to VAERS (the vaccine adverse events reporting system); the worrisome data that emerged during the vaccine trials (but were not disclosed in a forthcoming or honest fashion by the manufacturers); and other safety concerns that have been raised by thoughtful concerned scientists. [12-14]
For all of the above reasons we must not allow further vaccination of children against COVID until the COVID mass vaccination campaign has been thoroughly and properly re-evaluated. If the COVID mass vaccination campaign is thoroughly and honestly re-evaluated, there is no doubt in Dr. Vanden Bossche’s mind, or in my mind, that the COVID vaccines will be deemed unwise and unfit for human use.
As a pediatrician/pediatric rheumatologist with decades of experience, I strongly encourage pediatricians and parents to learn from Dr. Vanden Bossche and heed his warnings. In my view his voice is the most important, thoughtful, and scientifically accurate voice regarding the immunology, virology, vaccinology, and evolutionary biology of the COVID situation.

Note: The painting at the beginning of this article is “The Kiss” (Honore Daumier, 1808-1879). The above painting and the middle drawing are also by Daumier.
In addition to the References below, please see the APPENDIX OF MEDICAL ILLUSTRATIONS after the References.
REFERENCES:
- Vanden Bossche. Predictions on the Evolution of the COVID 19 Pandemic (Dr. Vanden Bossche):
https://www.voiceforscienceandsolidarity.org/scientific-blog/predictions-gvb-on-evolution-c-19-pandemic - Vanden Bossche. Immuno-epidemiologic ramifications of the C-19 mass vaccination experiment: Individual and global health consequences.
https://www.trialsitenews.com/a/immuno-epidemiologic-ramifications-of-the-c-19-mass-vaccination-experiment-individual-and-global-health-consequences.-1935ddcf - Vanden Bossche. The Immunologic Rationale Against C-19 Vaccination of Children. https://www.voiceforscienceandsolidarity.org/scientific-blog/the-immunological-rationale-against-c-19-vaccination-of-children
- Vanden Bossche, Rennebohm. Intra-pandemic vaccination of toddlers with non-replicating antibody-based vaccines targeted at ASLVI[1]– or ASLVD[2]-enabling glycosylated viruses prevents education of innate immune effector cells (NK cells).) https://www.trialsitenews.com/a/intra-pandemic-vaccination-of-toddlers-with-non-replicating-antibody-based-vaccines-targeted-at-aslvi1-or-aslvd2-enabling-glycosylated-viruses-pr-66e8b959
- Vanden Bossche, Rennebohm. Pediatricians, Internationally, Please Call for a Halt to the COVID Vaccination of Children. https://uploads-ssl.webflow.com/616004c52e87ed08692f5692/62e3848eb74bef65d5e602ac_COVID%20ANALYSIS%20%23111%20A%20CALL%20FOR%20A%20HALT_NO_link.pdf
- Vanden Bossche. Multisystem inflammatory syndrome in children (MIS-C) does NOT justify (at all!) their vaccination against SARS-CoV-2. https://uploads-ssl.webflow.com/616004c52e87ed08692f5692/62f0dea7d640657cfed14432_MIS-C%2Bfinal.pdf
- Vanden Bossche. Vaccination of vulnerable groups against monkeypox virus (MPV) in a highly C-19 vaccinated population will drive adaptive evolution of MPV and ignite multi-country epidemics in poorly C-19 vaccinated countries. https://uploads-ssl.webflow.com/616004c52e87ed08692f5692/62eccc023d3801059474162d_MPV%2Barticle.docx%2Bfinal(2).pdf
- Vanden Bossche. Novel bivalent C-19 vaccines: What does common immunological sense predict in regard to their impact on the C-19 pandemic? https://uploads-ssl.webflow.com/616004c52e87ed08692f5692/62ffa87af4dbe242b8425f6c_What%2Bto%2Bexpect%2Bfrom%2Bthe%2Bbivalent%2BC-19%2Bvaccines.pdf
- Dr. Vanden Bossche’s website: www.voiceforscienceandsolidarity.org
- Kimura, et al. Virological characteristics of the novel SARS-CoV-2 Omicron variants 2 including BA.2.12.1, BA.4 and BA.5 https://www.biorxiv.org/content/10.1101/2022.05.26.493539v1.full.pdf
- Uraki, et al. Characterization of SARS-CoV-2 Omicron BA.2.75 clinical isolates. https://www.biorxiv.org/content/10.1101/2022.08.26.505450v1
- Rennebohm. An Open Letter to Parents and Pediatricians Regarding COVID Vaccination. This is the original Open Letter. It provides 1078 references: https://notesfromthesocialclinic.org/an-open-letter-to-parents-and-pediatricians-2/
- Rennebohm, Vanden Bossche. Open Letter—Part II: A Review and Update https://notesfromthesocialclinic.org/open-letter-to-parents-and-pediatricians-part-ii-a-review-and-update/
- Rennebohm. Open Letter to Parents Regarding COVID Vaccination—Part III: Questions to Ask Your Physician—One Pediatrician’s Responses https://notesfromthesocialclinic.org/section-1-note-to-reader-table-of-contents/
- Rennebohm, McMillan: Video-interview regarding the initial Open Letter to Parents and Pediatricians: https://www.youtube.com/watch?v=uDRVq9NKrJQ&t=981s
- Reyneveld GI, Savelkoul HFJ and Parmentier HK (2020) Current Understanding of Natural Antibodies and Exploring the Possibilities of Modulation Using Veterinary Models. A Review. Front. Immunol. 11:2139. doi: 10.3389/fimmu.2020.02139
- Coutinho A, Kazatchkine MD, Avrameas S. Natural autoantibodies. Curr Opin Immunol. (1995) 7:812–8. doi: 10.1016/0952-7915(95)80053-0
- Holodick NE, Rodríguez-Zhurbenko N, Hernández AM. Defining natural antibodies. Front Immunol. (2017) 8:872. doi: 10.3389/fimmu.2017.00872
- Panda S, Ding JL. Natural antibodies bridge innate and adaptive immunity. J Immunol. (2015) 194:13–20. doi: 10.4049/jimmunol.1400844
- Palma, Joanna, et al. Natural antibodies–facts known and unknown. Central European Journal of Immunology 43.4 (2018): 466-475. https://www.termedia.pl/Natural-antibodies-facts-known-and-unknown,10,34697,0,1.html
- Ochsenbein AF, Zinkernagel RM. Natural antibodies and complement link innate and acquired immunity. Immunol Today. (2000) 21:624–30. doi: 10.1016/s0167-5699(00)01754-0
- Binder C. Naturally occurring IgM antibodies to oxidation-specific epitopes. Adv Exp Med Biol. (2012) 750:2–13. doi: 10.1007/978-1-4614-3461-0_1
- Carsetti, et al. The immune system of children: the key to understanding SARS-CoV-2 susceptibility? https://www.ncbi.nlm.nih.gov/pmc/articles/PMC7202830/
- Baumgarth, N., Tung, J.W. & Herzenberg, L.A. Inherent specificities in natural antibodies: a key to immune defense against pathogen invasion. Springer Semin Immun 26, 347–362 (2005).https://link.springer.com/article/10.1007/s00281-004-0182-2
- Prechl. A generalized quantitative antibody homeostasis model: antigen saturation, natural antibodies and a quantitative antibody network. Clinical & translational immunology 6.2 (2017): e131. https://onlinelibrary.wiley.com/doi/full/10.1038/cti.2016.90
- Abel, et al. Natural Killer cells: Development, Maturation, and clinical utilization. https://www.frontiersin.org/articles/10.3389/fimmu.2018.01869/full
- Orr, et al. Natural Killer Cell Education and Tolerance. https://www.sciencedirect.com/science/article/pii/S0092867410010007
- Arosh Shavinda Perera Molligoda Arachchige. Human NK cells: From development to effector functions. https://journals.sagepub.com/doi/pdf/10.1177/17534259211001512
- Nguyen, et al. Natural IgM prevents autoimmunity by enforcing B cell central tolerance induction. https://www.ncbi.nlm.nih.gov/labs/pmc/articles/PMC4324358/
- Netea, et al. Trained Immunity: a program of innate immune memory in health and disease. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5087274/pdf/nihms-824693.pdf
- Patel, et al. Immunological Outcomes of Antibody Binding to Glycans Shared Between Microorganisms and Mammals. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5119654/pdf/nihms810515.pdf
- Lau, et al. Epigenetic control of innate and adaptive immune memory. https://www.ncbi.nlm.nih.gov/labs/pmc/articles/PMC6225771/pdf/nihms-995099.pdf
- Iwasaki, et al. The potential danger of suboptimal antibody responses in COVID-19. https://www.nature.com/articles/s41577-020-0321-6.pdf?origin=ppub
- Wang, et al. Emerging vaccine-breakthrough SARS-CoV-2 variants. https://pubs.acs.org/doi/10.1021/acsinfecdis.1c00557
- Vanden Bossche. Poor virus-neutralizing capacity in highly C-19 vaccinated populations could soon lead to a fulminant spread of Sars- CoV-2 super variants that are highly infectious and highly virulent in vaccinees while being fully resistant to all existing and future spike-based C- 19 vaccines https://vitalhuman.substack.com/p/geert-vanden-bossches-analysis-of?s=r
- Vanden Bossche (2021) Why is the ongoing mass vaccination experiment driving a rapid evolutionary response of SARS-CoV-2? https://trialsitenews.com/why-is-the-ongoing-mass-vaccination-experiment-driving-a-rapidevolutionary-response-of-sars-cov-2/
- Sasaki, et al. Antigen escape selects for the evolution of higher pathogen transmission and virulence. https://www.nature.com/articles/s41559-021-01603-z
- Martin, et al. The emergence and ongoing convergent evolution of the N501Y lineages coincides with a major global shift in the SARS-CoV-2 selective landscape https://pubmed.ncbi.nlm.nih.gov/33688681/
- Vanden Bossche. https://www.voiceforscienceandsolidarity.org/scientific-blog/predictions-gvb-on-evolution-c-19-pandemic
- Van Egeren et al.: Risk of rapid evolutionary escape from biomedical interventions targeting SARS-CoV-2 spike protein https://pubmed.ncbi.nlm.nih.gov/33909660/
- Hoffmann, et al. The Omicron variant is highly resistant against antibody-mediated neutralization: Implications for control of the COVID-19 pandemic. https://pubmed.ncbi.nlm.nih.gov/35026151/
- Yahi, et al. Infection-enhancing anti- SARS-CoV-2 antibodies recognize both original Wuhan/D614G strain and Delta variant. A potential risk for mass vaccination: https://www.journalofinfection.com/article/S0163-4453(21)00392-3/fulltext
- Liu, et al. An infectivity-enhancing site on the SARS-CoV-2 spike protein targeted by antibodies. https://pubmed.nlm.nih.gov/33909660/
- Asarnow, et al. Structural insight into SARS-CoV-2 neutralizing antibodies and modulation of syncytia. https://pubmed.ncbi.nlm.nih.gov/33974910/
- Fantini, et al. Structural and molecular modelling studies reveal a new mechanism of action of chloroquine and hydroxychloroquine against SARS-CoV-2 infection. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC7128678/pdf/main.pdf
- Moretta, et al. NK-CTLs, a novel HLA-E-restricted T-cell subset. file:///C:/Users/rmren/Downloads/NK-CTLs,%20a%20novel%20HLA-E-restricted.pdf
- Chakradhar, Shraddha. Updated, augmented vaccines compete with original antigenic sin. Nature medicine 21.6 (2015): 540-542.
- Tripp, Ralph A., and Ultan F. Power. Original antigenic sin and respiratory syncytial virus vaccines. Vaccines 7.3 (2019): 107. https://www.mdpi.com/2076-393X/7/3/107/htm
- Rennebohm. A Call for an Independent International COVID Commission. https://notesfromthesocialclinic.org/a-call-for-an-independent-international-covid-commission/
Further references regarding the Education of NK Cells:
- Anfossi N, Andre P, Guia S, Falk CS, Roetynck S, Stewart CA, et al. Human NK cell education by inhibitory receptors for MHC class I. Immunity. (2006) 25:331–42. doi: 10.1016/j.immuni.2006.06.013
- Kim S, Poursine-Laurent J, Truscott SM, Lybarger L, Song YJ, Yang L, et al. Licensing of natural killer cells by host major histocompatibility complex class I molecules. Nature. (2005) 436:709–13. doi: 10.1038/nature03847
- Boudreau JE, Hsu KC. Natural killer cell education in human health and disease. Curr Opin Immunol. (2018) 50:102–11. doi: 10.1016/j.coi.2017.11.003
- Elliott JM, Yokoyama WM. Unifying concepts of MHC-dependent natural killer cell education. Trends Immunol. (2011) 32:364–72. doi: 10.1016/j.it.2011.06.001 163.
- Boudreau JE, Hsu KC. Natural killer cell education and the response to infection and cancer therapy: stay tuned. Trends Immunol. (2018) 39:222–39. doi: 10.1016/j.it.2017.12.001 164.
- Horowitz A, Djaoud Z, Nemat-Gorgani N, Blokhuis J, Hilton HG, Beziat V, et al. Class I HLA haplotypes form two schools that educate NK cells in different ways. Sci Immunol. (2016) 1:eaag1672. doi: 10.1126/sciimmunol.aag1672 165.
- Beziat V, Traherne JA, Liu LL, Jayaraman J, Enqvist M, Larsson S, et al. Influence of KIR gene copy number on natural killer cell education. Blood. (2013) 121:4703–7. doi: 10.1182/blood-2012-10-461442
- Fauriat C, Ivarsson MA, Ljunggren HG, Malmberg KJ, Michaelsson J. Education of human natural killer cells by activating killer cell immunoglobulin-like receptors. Blood. (2010) 115:1166–74. doi: 10.1182/blood-2009-09-245746
For additional articles on COVID, please see the following websites:
Dr. Geert Vanden Bossche’s Website: www.voiceforscienceandsolidarity.org
Dr. Rennebohm’s Website: www.notesfromthesocialclinic.org
APPENDIX OF MEDICAL ILLUSTRATIONS:
When a person is exposed to the SARS-CoV-2 virus, the virus typically enters through the person’s nose or throat and threatens to infect the person’s upper respiratory tract (URT). By “threatens to infect” we mean that the virus tries to enter the mucosal cells in the URT. The virus wants to enter those cells because it needs to replicate within our cells in order to survive. Viruses cannot replicate outside of living cells and, therefore, cannot survive outside of cells.
The SC-2 virus is able to enter cells because it has spike protein on its surface that it inserts into the ACE2 receptors on mucosal cells (and, potentially, other cells). When this insertion occurs, the ACE2 “door” of the cell opens and the virus enters the cell. The spike protein is the “key,” and the ACE2 receptor is the “keyhole.” Specifically, and more accurately, it is the Receptor Binding Domain (RBD) at the tip of the spike protein that is inserted into the “keyhole” to unlock the “door.”
Below are many illustrations that will help the reader to better visualize the appearance of the whole virus, the spike protein on the surface of the virus, the RBD at the tip of the spike protein, the ACE-2 receptor on human cells, the NTD (N-terminal domain) of the spike protein (where vaccinal non-neutralizing antibodies attach), and how antibodies bind to the RBD to block its attachment to the ACE-2 receptor.
For a review of how the immune system works, the reader is referred to the second section of the original Open Letter and to the entirety of the Open Letter—Part II.
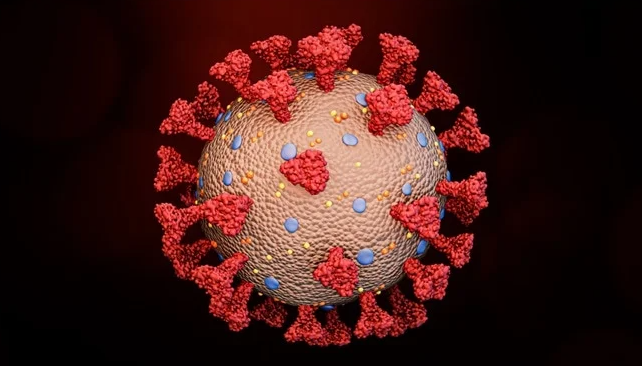
Figure 1: Here is the whole SARS-CoV-2 virus—the entire coronavirus, with many spike proteins (red) projecting from its surface.
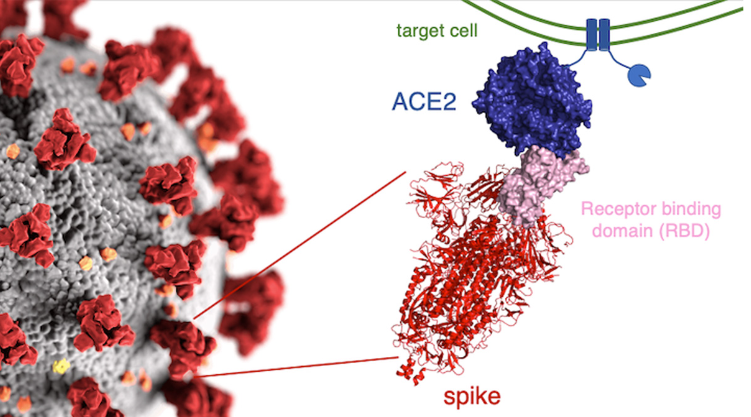
Figure 2 (above): On the left, many spike proteins (red) are sticking out from the surface of the whole virus. To the right of the whole virus is a drawing of what the actual spike protein looks like (in pink and red). At the tip of the spike protein is the Receptor Binding Domain (RBD), in pink. It is the RBD that fits into (or attaches to) the ACE2 receptor. The ACE2 receptor (in blue) is part of the wall of the human cell (the target cell) that the virus seeks to enter. When the RBD (the “key” ) fits into (attaches to) the ACE2 receptor (the “keyhole),” the door into the cell is opened and the virus enters the cell. The two little squiggly parts at the bottom of the spike protein are what anchor the spike protein into the surface of the virus.
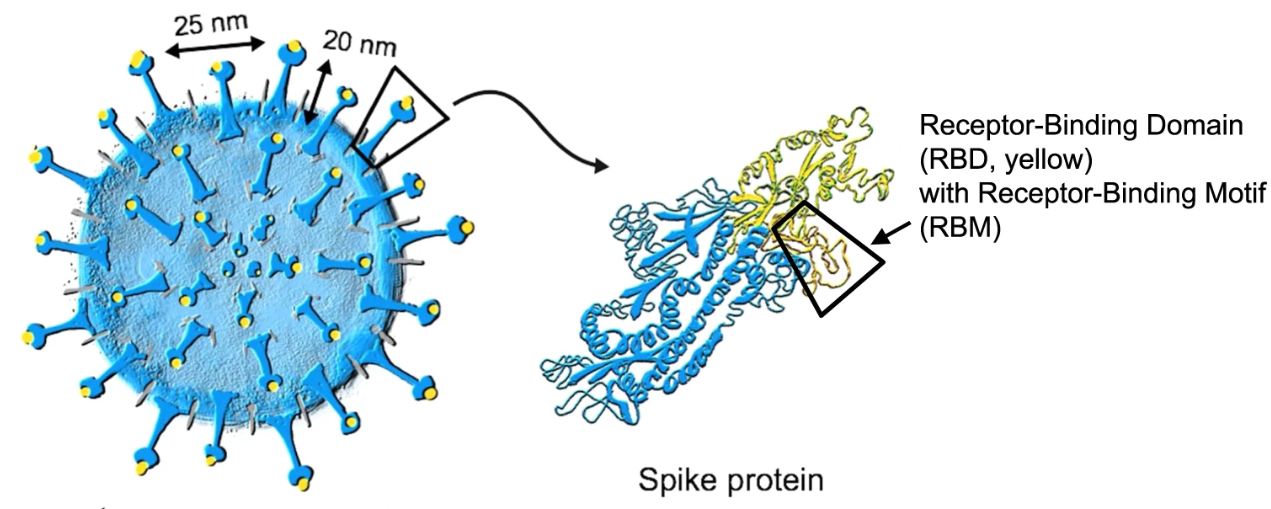
Figure 3: Another view of the whole virus, with its spike proteins. The RBD at the tip of the spike protein is shown in yellow.
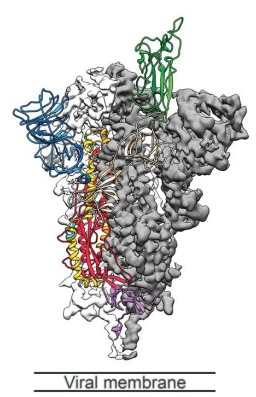
Figure 4: Another drawing of the spike protein. The RBD is shown in green (at the tip of the spike protein). The blue structure in the upper left is the N-terminal Domain (NTD). The gray area represents the lipid membrane of the virus.
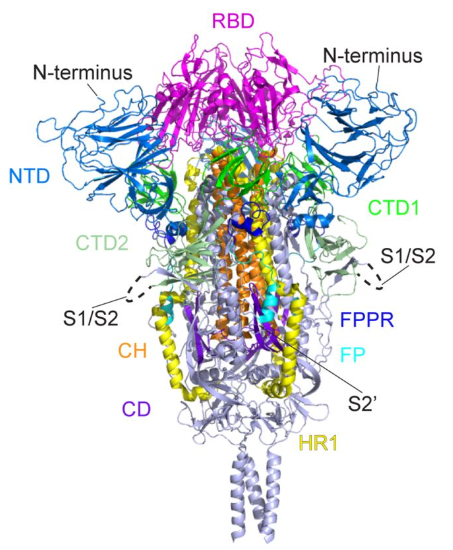
Figure 5: Another drawing of the spike protein. The RBD is at the tip (in magenta). There is an NTD region (in blue) to the left of the RBD and another NTD to the right of the RBD.
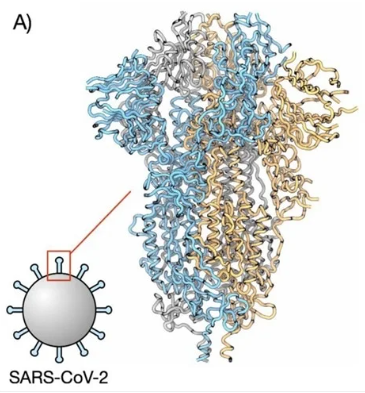
Figure 6: Another drawing of the spike protein.
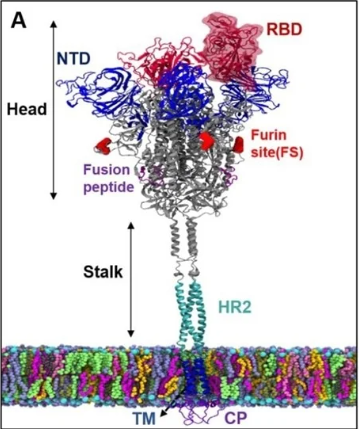
Figure 7: A spike protein anchored within the surface of the virus. The spike protein has a stalk and a head. At the tip of the head is the RBD. An NTD is also shown.
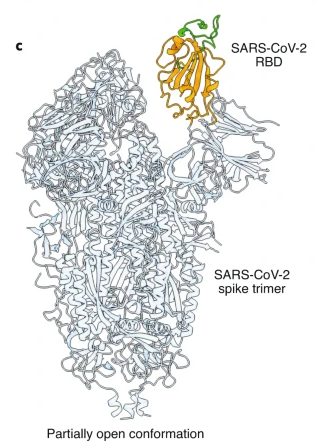
Figure 8: Another drawing of the spike protein. In this drawing the RBD is shown in its “open” position (conformation). When the RBD is in its “open” position, it more easily fits into (attaches to) the ACE2 “keyhole.”
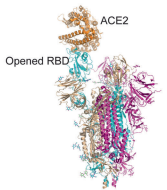
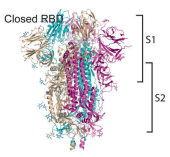
Figure 9: When the RBD is in the “open” position, it is better able to attach to the ACE2 receptor (which is on human cells). When the RBD is in the “closed” position, it is more difficult for it to attach to the ACE2 receptor. Note that the spike protein consists of an S1 region (the upper half) and an S2 region, the lower half. Both the RBD and NTD are in the S1 region.
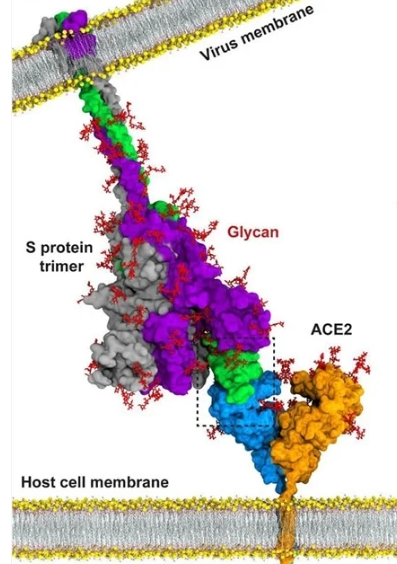
Figure 10: This is a drawing of the spike protein. It is anchored in the outside wall of the virus (the virus membrane). The RBD is shown in green, at the tip of the spike protein. The ACE2 receptor is anchored in the wall (membrane) of the human cell that the virus seeks to enter. The ACE2 receptor is shown with a blue half and a brown half. In this drawing, the RBD is firmly attached to the blue portion of the ACE2 receptor. This attachment will allow the virus to enter the human cell.
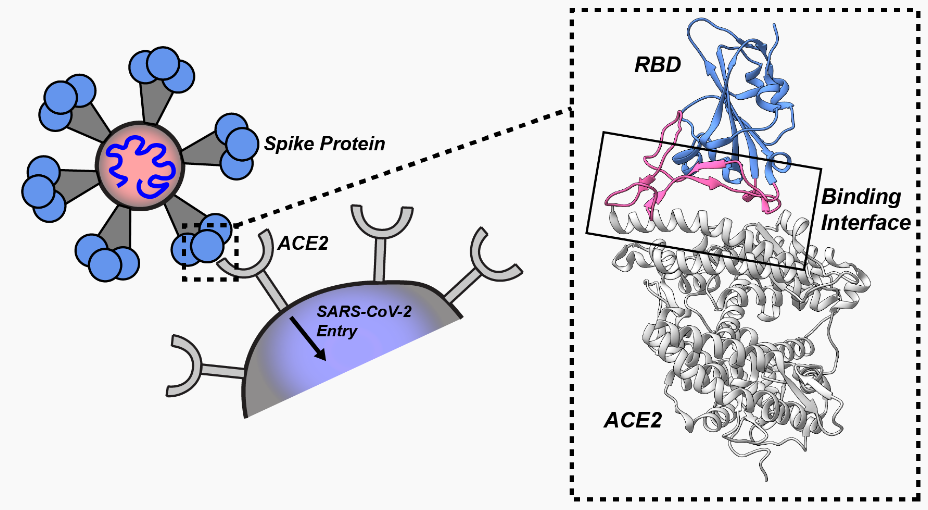
Figure 11: When the RBD (in blue) at the tip of the spike protein fits into (attaches to) the ACE2 receptor (which is on the surface of the human cell that the virus seeks to enter), that attachment results in the virus being able to enter the cell. In the dotted box you can see how the RBD attaches to the ACE2 receptor.
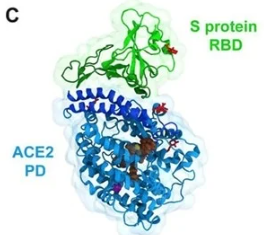
Figure 12: This drawing shows the RBD attached to the ACE2 receptor.
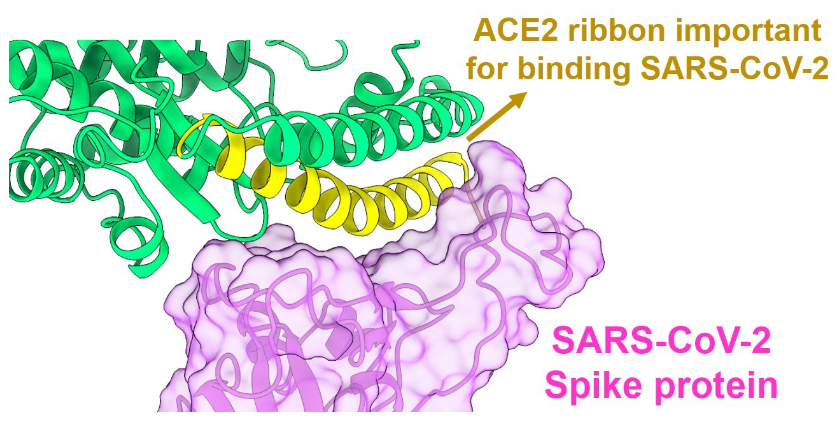
Figure 13: Another drawing of the RBD of the spike protein (in purple) successfully attached to (binding to) the ACE2 receptor (in green).
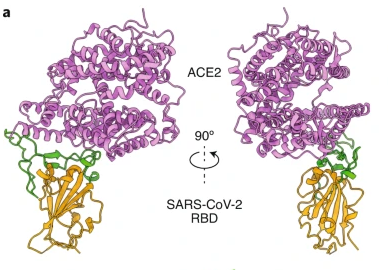
Figure 14: Another view of the RBD (brown and green) bound to the ACE2 receptor (purple).
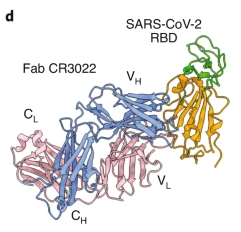
Figure 15: In this drawing a neutralizing antibody (FAB CR3022, blue and pink) has attached to the RBD (brown and green). When an antibody is firmly and tightly attached to the RBD, the RBD will be unable to attach to an ACE2 receptor—i.e., the antibody is neutralizing. The neutralizing antibody blocks the attachment of the RBD to the ACE2 receptor.
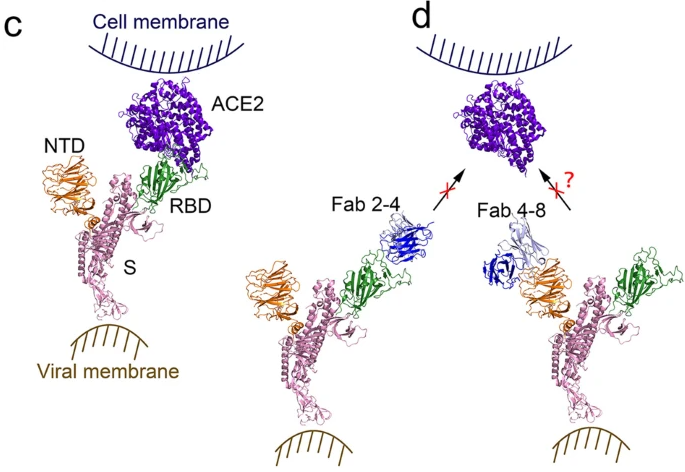
Figure 16: On the left (image C), the RBD at the tip of the spike protein has successfully bound to the ACE2 receptor on the human cell. On the right (image d, note that a neutralizing antibody (Fab 2-4, in blue) has attached to the RBD (in green), and this blocks (prevents) that RBD from attaching to the ACE2 receptor (in purple). Consequently, that virus cannot enter the human cell. On the far right, an antibody (Fab 4-8, in blue) has attached to the NTD (N-terminal domain, in brown) of the spike protein. Early during the pandemic it was suspected that some antibodies to NTD might also somehow block attachment of spike protein to ACE2 receptor—but this was unclear, questionable. Recent studies have demonstrated that, in the absence of effective neutralizing antibodies to the RBD, some antibodies that attach to the NTD can cause conformational changes in the spike protein that result in the RBD moving into an “open” position, and this facilitates RBD attachment to the ACE2 receptor and entry of virus into the human cell. It is now realized that there are antibodies to the NTD that are non-neutralizing antibodies, and that, in the absence of effective neutralizing antibodies, these non-neutralizing antibodies actually facilitate entry of virus into human cells. They do not block entry of virus into cells, they facilitate entry. The COVID vaccines result in production of large quantities of neutralizing antibodies (to the RBD) and non-neutralizing antibodies (to the NTD). When the vaccinal neutralizing antibodies of vaccinated individuals cease to match the mutated RBD of the spike protein of new variants, there is concern that vaccinal non-neutralizing antibodies (to NTD) are able to bind more tightly to the NTD and may facilitate entry of virus into cells. This is why vaccinated individuals may be more easily infected by the Omicron variants that have become resistant to the vaccinal neutralizing antibodies.
Figure 17: This is a drawing of a neutralizing antibody to the RBD of the spike protein. This antibody will block (prevent) attachment of the RBD to the ACE2 receptor.
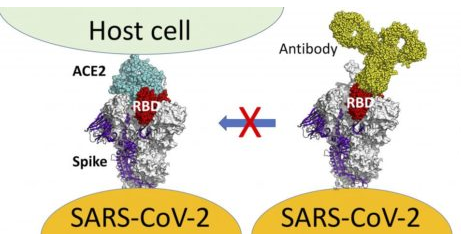
Figure 18: On the left, the RBD at the tip of the spike protein of the virus has successfully and firmly attached to the ACE2 receptor on the human cell. On the right, a neutralizing antibody to the RBD has firmly attached to the RBD. This antibody will prevent the RBD from attaching to the ACE2 receptor.
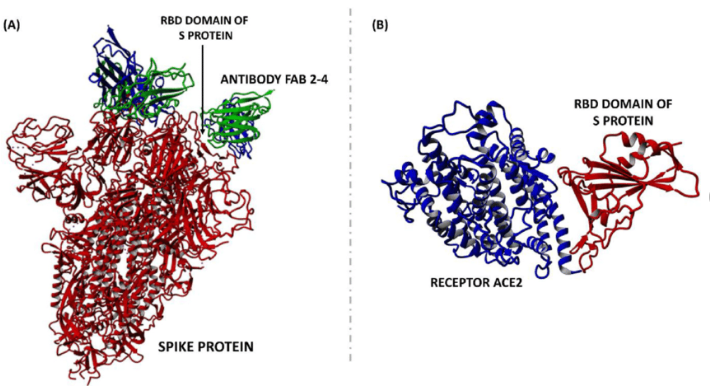
Figure 19: On the left, a neutralizing antibody to RBD (FAB 2-4) has attached to the RBD of the spike protein. This antibody will prevent attachment of the RBD to the ACE2 receptor on human cells. On the right is the usual attachment of RBD to ACE2 that is prevented by neutralizing antibody.
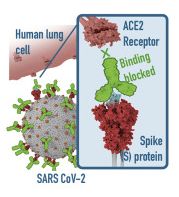
Figure 20: In this drawing, neutralizing antibodies have attached to the RBD of the spike proteins. This blocks attachment of the RBD to ACE2 receptors on human cells. That is how neutralizing antibodies prevent virus from entering human cells.
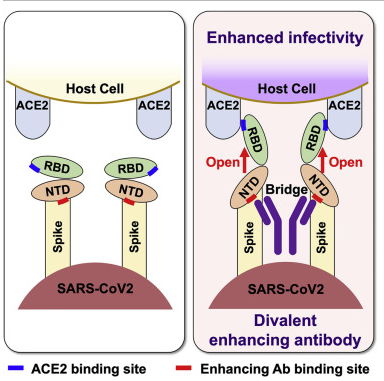
Figure 21: The drawing on the left shows the spike protein (with its RBD and NTD) approaching a human cell. The drawing on the right shows a non-neutralizing antibody (the Y-shaped structure in dark purple) that is attached to the two NTDs. When this non-neutralizing antibody attaches to the NTD, it flips the RBD into the “open” position, which is the ideal position for the RBD to successfully achieve attachment to the ACE2 receptor. In this way, the non-neutralizing antibodies to the NTD actually enhance entry of the virus into the human cell—i.e., increases infectivity.
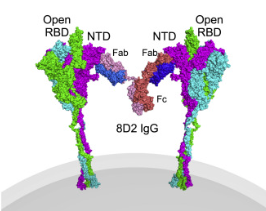
Figure 22: Another drawing of a non-neutralizing antibody (the Y-shaped structure consisting of two Fab fragments and an Fc stalk) that is attached to the NTD of two neighboring spike proteins, resulting in flipping the RBD into the “open” position, thereby enhancing infectivity (viral entry into the human cell).
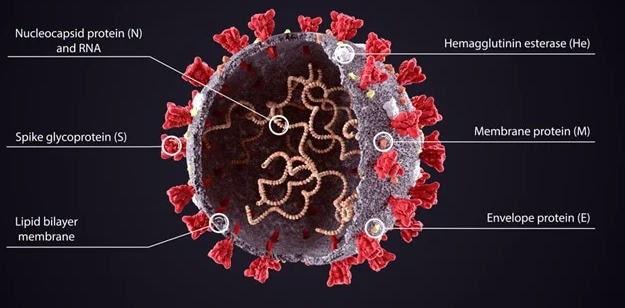
Figure 23: This is the SARS-CoV-2 (SC-2) virus, covered with its spike proteins. Note that there are several other important proteins on and within the virus, besides the spike protein: Nucleocapsid (N) protein, Membrane (M) protein, Envelope (E) protein. The normal, natural immune system wisely produces antibodies not only to the spike protein, but also to many other proteins on/in the virus—including antibodies to nucleocapsid protein. The COVID vaccines train the immune system to produce neutralizing and non-neutralizing antibodies to the spike protein, but not to nucleocapsid protein or other proteins in/on the virus.
Lab testing of people with naturally acquired immunity will reveal antibodies to spike protein and to nucleocapsid protein. Lab testing of people who have been vaccinated and have never had actual natural infection, will reveal anti-spike antibodies but not antibodies to nucleoprotein.
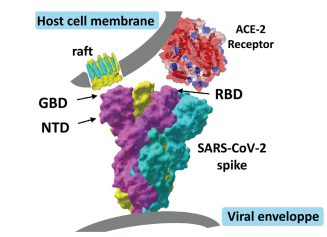
Figure 24: How might hydroxychloroquine inhibit viral entry into human cells? Viral entry requires dual recognition of gangliosides and angiotensin-converting enzyme-2 (ACE-2) by the SARS-CoV-2 spike (S) protein. The viral protein displays two distinct domains, the tips of which are available for distinct types of interactions. The receptor-binding domain (RBD) binds to the ACE-2 receptor, and the N-terminal domain (NTD) binds to the ganglioside-rich domain of the plasma membrane. Lipid rafts, which are membrane domains enriched in gangliosides (in yellow) and cholesterol (in blue), provide a perfect attractive interface for adequately positioning the viral S protein at the first step of the infection process. Structural and molecular modelling studies suggest that amino acid residues 111–162 of the NTD form a functional ganglioside-binding domain (GBD), the interaction of which with lipid rafts can be efficiently prevented by chloroquine and hydroxychloroquine. See Figure 25.
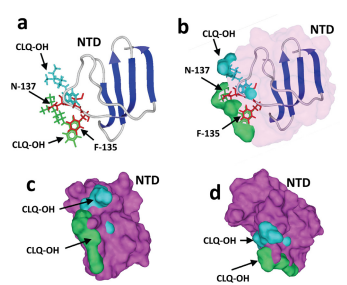
Figure 25: Mechanism of action of hydroxychloroquine (CLQ-OH). The N-terminal domain (NTD) bound to GM1 was superposed onto GM1 in interaction with two CLQOH molecules. The models only show the NTD and both CLQ-OH molecules (not GM1, to improve clarity). (a, b) The aromatic ring of F-135 (in red), which stacks onto the glucose cycle of GM1, overlaps the aromatic CLQ-OH rings (in green) which also bind to GM1 via a CH-π stacking mechanism. The N-137 residue of the NTD interacts with the N-acetylgalactosamine residue of GM1, but this interaction cannot occur in the presence of CLQ-OH as this part of GM1 is totally masked by the drug. (c, d) Superposition of the NTD surface (in purple) with the two CLQ-OH molecules bound to GM1, illustrating the steric impossibility that prevents NTD binding to GM1 when both CLQ-OH molecules are already interacting with the ganglioside.
NOTE RE: SOURCES OF MEDICAL ILLUSTRATIONS:
The above Figures 21 and 22 may be found in an article by Liu, et al. at: https://www.sciencedirect.com/science/article/pii/S0092867421006620
Figures 24 and 25 may be found in an article by Fantini, et al: Structural and molecular modelling studies reveal a new mechanism of action of chloroquine and hydroxychloroquine against SARS-CoV-2 infection. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC7128678/pdf/main.pdf
All of the other Figures/images (and their sources) may be found by searching for “spike protein N-terminal domain” at:
0 Comments